
Chromista
règne d'organismes eucaryotes
From Wikipedia, the free encyclopedia
La taxinomie des Chromista est en pleine évolution voire révolution, et les différentes classifications sont très disparates, notamment entre les classifications qui maintiennent une taxonomie plus classique et les classifications purement cladistiques qui ont réparti le groupe dans les clades SAR, Telonemia (en), Haptista, Pancryptista (en).

| Clade | Biota |
|---|---|
| Clade | Neomura |
| Domaine | Eukaryota |
| Clade | Neokaryotes |
| Clade | Neozoa |
| Super-règne | Corticata |
Sous-règnes de rang inférieur
Les Chromistes[a] (Chromista) sont un règne d'organismes uni- et pluricellulaires eucaryotes regroupant des taxons historiquement considérés comme des protozoaires (Foraminifères, Radiolaires, Ciliés, etc.), comme des champignons (Oomycètes, Labyrinthulomycètes, etc.) et des algues (Algues brunes, Diatomées, Dinoflagellés, Coccolithophores, etc.).
Historique
Ce règne a été proposé par le biologiste britannique Thomas Cavalier-Smith en 1981[1]. Les Chromista étaient d'abord un clade supposé d'organismes eucaryotes pour la plupart photosynthétiques, regroupant également certains organismes autrefois classés parmi les « champignons inférieurs » — donc hétérotrophes. La pertinence de ce clade a été remise en cause par des études de 2007[2] rapprochant les Stramenopiles des Rhizaria. Il a donc été proposé d'utiliser le terme de Chromista pour désigner l'ensemble présumé monophylétique formé par les Harosa et les Hacrobia à la place du terme de Chromalveolata au sens large[3].
Les Chromistes sont moins diversifiés que les algues vertes mais plus que les algues rouges[4].
Le groupe Chromista est polyphylétique, alors, ce nom n'est plus utilisé pour le groupe[5],[6],[7].
Étymologie
Le terme Chromista dérive du grec χρῶμα / chróma, couleur, en référence aux pigments de ces organismes qui leur donne une teinte plus ou moins soutenue. Le terme fut inventé en 1981 par le biologiste britannique Thomas Cavalier-Smith, sur la base de termes plus anciens, par exemple Chromophyta.
Caractéristiques propres
On trouve des caractéristiques propres au groupe, qui peuvent avoir disparu secondairement :
- Un réticulum périplastidial, situé entre les deux enveloppes du chloroplaste et les deux membranes externes.
- Les deux sous-groupes Straménopiles et Haptophytes présentent les mêmes pigments photosynthétiques.
- Le chloroplaste de la cellule est constitué de quatre membranes : les deux membranes communément trouvées chez les chloroplastes et deux membranes externes reliées le plus souvent à l'enveloppe nucléaire. Cette situation serait le résultat d'une endosymbiose secondaire et plus précisément d'une ingestion d'une Rhodophyte unicellulaire par un autre Eucaryote. À ce sujet, la communauté scientifique cherche à savoir si l'endosymbiose s'est effectuée plusieurs fois au sein des Chromistes ou si cela a été réalisé une seule fois — et l'endosymbiote ensuite perdu —, ce qui résoudrait le problème des Straménopiles non photosynthétiques.
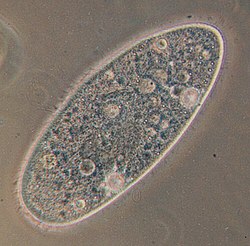

 Plasmodium falciparum (Alveolata, Apicomplexa), parasite responsable du paludisme (stade en anneau dans une hématie).
Plasmodium falciparum (Alveolata, Apicomplexa), parasite responsable du paludisme (stade en anneau dans une hématie).








Géologie
Le plus ancien fossile connu présumé date du Précambrien (700 Ma).
En effet, le Précambrien récent a livré des fossiles de type « film carboné », se présentant principalement sous la forme de simples empreintes noirâtres dans la roche. Si certaines de ces compressions sont dépourvues de caractéristiques distinctes et correspondraient à d'anciens tapis microbiens, d'autres exhibent une structure dotée d'un thalle. Cette morphologie suggère qu'elles appartiennent à des organismes analogues à des algues, peut-être des algues brunes, ou phéophytes. Bien que les phéophytes puissent avoir des origines remontant à l'Ordovicien, leurs traces fossiles demeurent rares avant le Miocène[8],[9].
Les chromistes dotés d'un squelette minéral, de silice ou de carbonate de calcium, possèdent un registre fossile bien plus fourni. L'accumulation de leurs tests fossilisés, comme ceux des diatomées, des coccolithophores et des silicoflagellés, est parfois à l'origine de formations rocheuses entières. Leur origine est ancienne, avec des fossiles potentiels mais contestés dès le Précambrien récent. Les premiers diatomées et coccolithophores incontestables apparaissent au Jurassique, tandis que les chrysophytes et les silicoflagellés sont avérés à partir du Crétacé. Pour la majorité des autres chromistes, les témoignages fossiles, extrêmement fragmentaires, ne remontent pas au-delà de cette même période. Au Cénozoïque, les restes de chromistes siliceux, notamment les diatomées et les chrysophytes, deviennent souvent prépondérants dans les sédiments lacustres[8].
Liste des embranchements/divisions
Selon AlgaeBase (29 janvier 2022) :
- Embranchement des Bacillariophyta Karsten
- Embranchement des Bigyra Cavalier-Smith
- Embranchement des Cryptophyta Cavalier-Smith
- Embranchement des Katablepharidophyta N.Okamoto & I.Inouye
- Embranchement des Ochrophyta Cavalier-Smith
- Embranchement des Telonemia Shalchian-Tabrizi & al.
- Embranchement des Chromista phylum incertae sedis
- sous-règne des Hacrobia
- Embranchement des Cryptista Cavalier-Smith
- Embranchement des Haptophyta Hibberd ex Edvardsen & Eikrem
- sous-règne des Harosa (supergroupe SAR)
- infra-règne des Halvaria (ou Heterokonta ?)
- Embranchement des Ciliophora Doflein
- Embranchement des Miozoa Cavalier-Smith
- Embranchement des Oomycota Arx
- infra-règne des Rhizaria
- Embranchement des Foraminiferida d'Orbigny
- infra-règne des Halvaria (ou Heterokonta ?)
Selon Cavalier-Smith (1981, 2004, 2010)[10],[11],[3] :
- sous-règne des Harosa (Cavalier-Smith 2010)
- infrarègne des Heterokonta (Cavalier-Smith 1986)
- embranchement des Bigyra (Cavalier-Smith 1998)
- embranchement des Pseudofungi (Cavalier-Smith 1986)
- embranchement des Ochrophyta (Cavalier-Smith 1986)
- infrarègne des Alveolata (Cavalier-Smith 1991)
- embranchement des Myzozoa (Cavalier-Smith 2004)
- sous-embranchement des Dinozoa (Cavalier-Smith 1981)
- sous-embranchement des Apicomplexa (Levine 1970)
- embranchement des Ciliophora (Doflein 1901)
- embranchement des Myzozoa (Cavalier-Smith 2004)
- infrarègne des Rhizaria (Cavalier-Smith 2002)
- embranchement des Cercozoa (Cavalier-Smith 1998)
- embranchement des Retaria (Cavalier-Smith 1999)
- sous-embranchement des Foraminifera (Eichwald 1830)
- sous-embranchement des Radiozoa (Cavalier-Smith 1987)
- infrarègne des Heterokonta (Cavalier-Smith 1986)
- sous-règne des Hacrobia (Okamoto et al. 2009)
- embranchement des Cryptista (Cavalier-Smith 1989)
- embranchement des Haptophyta (Hibberd 1976)
- embranchement des Heliozoa (Haeckel 1862)
Selon ITIS (13 mars 2012)[12] :
- division des Bacillariophyta
Selon Catalogue of Life (13 mars 2012)[13] :
- embranchement des Cryptophyta
- embranchement des Haptophyta
- embranchement des Hyphochytriomycota
- embranchement des Labyrinthista
- embranchement des Ochrophyta
- embranchement des Oomycota
- embranchement des Sagenista (en)
- embranchement des Chromista phylum incertae sedis
Selon World Register of Marine Species (13 mars 2012)[14] :
- sous-règne Hacrobia
- embranchement des Cryptophyta
- embranchement des Haptophyta
- embranchement des Heliozoa
- sous-règne des Harosa
- infra-règne des Alveolata
- embranchement des Ciliophora
- embranchement des Myzozoa
- infra-règne des Heterokonta
- embranchement des Bigyra
- embranchement des Ochrophyta
- embranchement des Oomycota
- infra-règne des Rhizaria
- embranchement des Cercozoa
- embranchement des Foraminifera
- embranchement des Radiozoa
- infra-règne des Alveolata
Selon Aubert (2017)[15] :
- imperium Eukaryota
- regnum Chromista
- subregnum Hacrobia
- subregnum Harosa
- infraregnum Rhizaria
- infraregnum Halvaria
- superphylum Alveolata
- phylum Myzozoa
- phylum Ciliophora
- superphylum Heterokonta
- phylum Bigyra
- phylum Pseudofungi
- phylum Ochrophyta
- superphylum Alveolata
- regnum Chromista
Voir aussi
Articles connexes
Liens externes
- (en) AlgaeBase : règne Chromista (+classification) (consulté le )
- (en) BioLib : Chromista Cavalier-Smith, 1981 (consulté le )
- (en) Catalogue of Life : Chromista (consulté le )
- (fr + en) GBIF : Chromista (consulté le )
- (en) IRMNG : Chromista (consulté le )
- (fr + en) ITIS : Chromista (consulté le )
- (en) OEPP : Chromista (consulté le )
- (en) Paleobiology Database : Chromista (consulté le )
- (en) Taxonomicon : Chromista Caval.-Sm. (1981) (consulté le )
- (en) UICN : taxon Chromista (consulté le )
- (en) WoRMS : Chromista (+ liste embranchements + liste classes) (consulté le )
- Chromista (Univ Paris Sud 11)
- (en) UCMP - University of California Museum of Paleontology, « Introduction to the Chromista » (consulté le )