Virus de la grippe
From Wikipedia, the free encyclopedia

Les virus de la grippe sont un ensemble de quatre espèces de virus à ARN monocaténaire de polarité négative distinguées chacune par un type antigénique particulier : le virus de la grippe A, le virus de la grippe B, le virus de la grippe C et le virus de la grippe D[1]. Parmi ces quatre types antigéniques, le type A est le plus dangereux, le type B présente moins de risques mais reste susceptible de provoquer des épidémies, et le type C n'est généralement associé qu'à des symptômes mineurs. Le type D est moins répandu que les autres et n'est pas connu pour provoquer des infections chez l'homme. Ces quatre virus ont un génome segmenté, à huit segments pour les deux premiers et à sept segments pour les deux derniers. La composition en acides aminés des virus de la grippe C et D est semblable à 50 %, taux semblable à celui observé entre les virus de la grippe A et B ; le taux de divergence entre les virus A et B d'une part et les virus C et D d'autre part est en revanche bien plus élevé[2].
Les antigènes, protéine de matrice (M1) et nucléoprotéine (NP), sont utilisés pour déterminer si un virus de la grippe est de type A, B, C ou D. La protéine M1 est nécessaire pour l'assemblage du virus et la nucléoprotéine intervient dans la transcription et la réplication virale[3],[4].
Des glycoprotéines se trouvent également à la surface de la membrane cellulaire. Les virus de la grippe A et B ont deux glycoprotéines : l'hémagglutinine (HA) et la neuraminidase (NA) ; les virus de la grippe C et D n'ont qu'une glycoprotéine, la glycoprotéine de fusion hémagglutinine-estérase (HEF)[5]. Ces glycoprotéines permettent la fixation et la fusion entre l'enveloppe virale et la membrane cellulaire. La fusion de ces membranes permet aux protéines et au génome du virus d'être libérés dans la cellule hôte, ce qui provoque alors l'infection[6]. Les types C et D sont les seuls virus de la grippe à exprimer une estérase. Cette enzyme est semblable à la neuraminidase produite par les virus de la grippe A et B en ce que ces deux enzymes détruisent les récepteurs des cellules hôtes.
Les glycoprotéines peuvent subir des mutations (glissement antigénique) ou un réassortiment dans lesquels une nouvelle hémagglutinine ou une nouvelle neuraminidase sont produites (cassure antigénique). Les virus de la grippe C et D ne peuvent donner lieu qu'à un glissement antigénique, alors que le virus de la grippe A connaît également des cassures antigéniques. Lorsque l'un ou l'autre de ces processus se produit, les anticorps produits par le système immunitaire de l'hôte ne protègent plus contre ces glycoprotéines modifiées. Pour cette raison, ces virus provoquent continuellement des infections.
Virus de la grippe A
Les quatre types de virus de la grippe correspondent chacun à une espèce de virus au sens de l'ICTV[7]. Chacune de ces espèces est à son tour subdivisée en sous-types, en sérotypes et en souches, dont la virulence et l'infectivité (en) varient sensiblement d'une espèce hôte à une autre.
| Types | Hôtes | Sous-types |
|---|---|---|
| Virus de la grippe A | Humains, porcs, oiseaux, chevaux, chauves-souris | H1N1, H1N2, H2N2, H3N1, H3N2, H3N8, H5N1, H5N2, H5N3, H5N8, H5N9, H7N1, H7N2, H7N3, H7N4, H7N7, H7N9, H9N2, H10N7 |
| Virus de la grippe B | Humains, phoques | Victoria, Yamagata |
| Virus de la grippe C | Humains, porcs, chiens | |
| Virus de la grippe D | Porcs, bovins |
Les virus de la grippe A sont classés en fonction des glycoprotéines de leur enveloppe virale, l'hémagglutinine et la neuraminidase, ce qui identifiée des sous-types notés HxNy, où H identifie l'hémagglutinine et N identifie la neuraminidase, avec x variant de 1 à 18 et y variant de 1 à 11. Ce sont les plus virulents de virus de la grippe, les suivants ayant été observés chez les humains :
- le sous-type H1N1, à l'origine de l'épidémie de « grippe espagnole » de 1918, de « grippe russe » de 1977 et de « grippe porcine » de 2009[8] ;
- le sous-type H2N2, à l'origine de l'épidémie de « grippe asiatique » de 1956-1958 ;
- le sous-type H3N2, à l'origine de la « grippe de Hong Kong » de 1968-1969 ;
- le sous-type H5N1, à l'origine de la « grippe aviaire » de 2006, et considéré comme une menace de pandémie[9] ;
- le sous-type H7N7, qui présente un risque zoonotique particulièrement élevé[10] ;
- les sous-types H7N2, H7N3[11], H9N2 et H10N7[12] ont été sporadiquement observés chez des humains ;
- le sous-type H3N8 a pu être impliqué dans l'épidémie de 1889-1890[13], qui a tué plus d'un million de personnes.
Au sein de ces sous-types, on distingue également des souches et des variantes, identifiées selon une nomenclature indiquant respectivement le type, l'origine géographique, le numéro de la souche et l'année d'identification, par exemple :
- A/Fujian/411/2002 (H3N2) ;
- A/Brisbane/59/2007 (H1N1).
Les souches à l'origine de l'épidémie de « grippe aviaire » de 2006 sont généralement désignées par HPAI H5N1, signifiant Highly Pathogenic Avian Influenza[14],[15].
| Sous-types de virus de la grippe A | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H | N | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 |
| 1 | Grippe espagnole-1918
Grippe russe-1977 Grippe porcine-2009 |
|||||||||||
| 2 | Grippe asiatique 1956 | |||||||||||
| 3 | Grippe de Hong-Kong | Grippe de 1889 | ||||||||||
| 4 | ||||||||||||
| 5 | GA | GA | GA | |||||||||
| 6 | ||||||||||||
| 7 | GA | GA | GA | GA | GA | |||||||
| 8 | ||||||||||||
| 9 | GA | |||||||||||
| 10 | ||||||||||||
| 11 | ||||||||||||
| 12 | ||||||||||||
| 13 | ||||||||||||
| 14 | ||||||||||||
| 15 | ||||||||||||
| 16 | ||||||||||||
| 17 | ||||||||||||
| 18 | ||||||||||||
| Grippe aviaire transmisse à l' homme avec décès | ||||||||||||
| Grippe aviaire transmisse à l' homme sans décès | ||||||||||||
Virus de la grippe B
Le virus de la grippe B est presque exclusivement humain, et moins répandu que le virus de la grippe A. Le seul hôte non humain connu pour héberger ce virus est le phoque[16]. Le virus de la grippe B mute deux à trois fois moins vite que celui de type A[17] et présente donc une variabilité génétique plus faible, avec un seul sérotype[18]. Pour cette raison, l'immunité contre les virus de la grippe B est généralement atteinte à un âge assez jeune. Le taux de mutation n'est cependant pas nul, et deux sous-types de virus de la grippe B sont clairement identifiés, appelés Victoria et Yamagata[19], de sorte que l'immunité ne peut être totale ni permanente. Néanmoins, l'absence d'hôtes non humains écarte la possibilité de l'émergence d'une pandémie à virus de grippe B[20]. Les variantes de ce virus sont identifiées par une nomenclature semblable à celle du virus de la grippe A, par exemple :
- B/Victoria/504/2000 ;
- B/Yamagata/16/88.
Virus de la grippe C
Le virus de la grippe C infecte les humains et les porcs, et est susceptible de provoquer des affections sévères ainsi que des épidémies localisées[21]. Ce type de virus de la grippe est cependant moins courant que les autres et semble provoquer essentiellement des affections bénignes chez l'enfant[22],[23].
Virus de la grippe D
Les premiers virus de la grippe D ont été isolés en 2011[24]. Ce type semble avoir divergé des virus de la grippe C il y a plusieurs centaines d'années[25]. On en connaît au moins deux souches[26]. Il semble infecter avant tout les bovins, mais a également été observé chez le porc.
Génome et structure
Les virus de la grippe appartiennent à l'ordre des Articulavirales, de sorte qu'ils présentent un génome segmenté, c'est-à-dire divisé en molécules d'acides nucléiques distinctes, en l'occurrence de sept à huit segments d'ARN monocaténaire de polarité négative. La longueur totale de ce génome est comprise entre 12 000 et 15 000 nucléotides, avec, par exemple pour le virus de la grippe A (H1N1), une distribution du type :
| Segment d'ARN | Protéine(s) | Nucléotides | Acides aminés |
|---|---|---|---|
| PB1 | Polymérase basique 1 | 2300 à 2500 | 757 + 87 (F2) |
| PB2 | Polymérase basique 2 | 2300 à 2500 | 759 |
| PA | Polymérase acide | 2200 à 2300 | 716 |
| HA | Hémagglutinine | 1700 à 1800 | 550 |
| NP | Nucléoprotéine | 1500 à 1600 | 498 |
| NA | Neuraminidase | 1400 à 1500 | 454 |
| M | Protéines membranaires | 1000 à 1100 | 252 + 97 |
| NS | Protéines non structurelles | 800 à 900 | 230 + 121 |
Parmi les segments ci-dessus, les deux plus importants sont PB1 et HA. PB1 donne une ARN polymérase ARN-dépendante essentielle à la virulence du virus, tandis que HA donne l'antigène glycoprotéique de surface (l'hémagglutinine) déterminant sa transmissibilité.
La séquence du génome présente des séquences terminales répétées à chaque extrémité. Les séquences répétées à l'extrémité 5' ont une longueur de 12 ou 13 nucléotides, celles de l'extrémité 3' ont une longueur de 9 à 11 nucléotides. Le virus de la grippe A produit une protéine PB1-F2 à partir d'un cadre de lecture ouvert alternatif du segment PB1, tandis que les segments M et NS produisent deux protéines chacun, notées M1, M2, NS1 et NS2, par épissage alternatif[27].
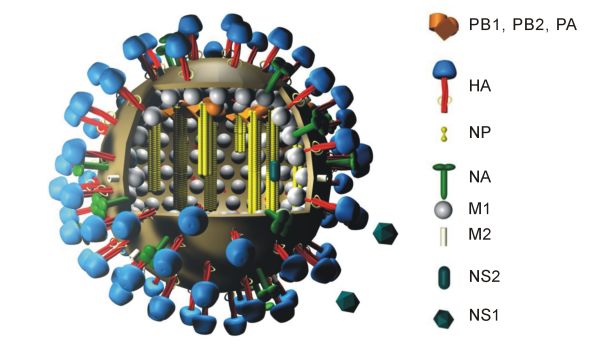 Structure générique d'un virus de la grippe A indiquant l'agencement de 10 protéines virales. La protéine PB1-F2 et les huit segments d'ARN ne sont pas représentés sur ce schéma.
Structure générique d'un virus de la grippe A indiquant l'agencement de 10 protéines virales. La protéine PB1-F2 et les huit segments d'ARN ne sont pas représentés sur ce schéma.

Les virions du virus de la grippe A ont un diamètre de 80 à 120 nm et forment généralement un ensemble de particules à la fois ellipsoïdales, bacilliformes et filamenteuses[28],[29],[30]. Leur génome a une longueur totale de 13,5 kilobases distribuées sur huit segments d'ARN monocaténaire de polarité négative qui codent 11 protéines, appelées HA, NA, NP, M1, M2, NS1, NEP, PA, PB1, PB1-F2 et PB2[31]. L'hémagglutinine HA et la neuraminidase NA sont les mieux caractérisées d'entre elles. Ce sont de grosses glycoprotéines situées sur la surface des virions. La neuraminidase est une enzyme intervenant dans le bourgeonnement des virions nouvellement formés dans les cellules infectées, tandis que l'hémagglutinine est une lectine intervenant dans la liaison du virus à la cellule hôte et dans l'injection du génome viral dans cette dernière[32]. L'hémagglutinine et la neuraminidase sont les cibles des antiviraux[33]. Ces protéines sont des antigènes reconnus par les anticorps de l'organisme[34].
.jpg)
Les virions sont pléomorphes, avec une enveloppe généralement ellipsoïde de 80 à 120 nm de diamètre et peuvent former des filaments atteignant 20 μm de long[29]. L'enveloppe est hérissée d'environ 500 protubérances faisant saillie d'environ 10 à 14 nm hors de la surface du virus. Les molécules d'hémagglutinine sont séparées les unes des autres par des grappes de neuraminidase, avec un ratio d'environ 4 à 5 molécules de neuraminidase par molécule d'hémagglutinine.
La membrane virale, chargée de cholestérol et hérissée de glycoprotéines, renferme la nucléocapside, qui contient des nucléoprotéines de différentes tailles arrangées selon une disposition mal connue. Les protéines ribonucléaires sont filamenteuses, avec un diamètre de 9 à 15 nm et une longueur de 50 à 130 nm. Elles présentent une symétrie hélicoïdale.
Cycle de réplication
Les mammifères infectés transmettent la grippe généralement par voie aérienne, par la toux ou ses éternuements expulsant des aérosols contenant le virus, tandis que les oiseaux contaminés la transmettent par leurs excréments. La grippe peut également être transmise par la salive, les sécrétions nasales, les selles et le sang. Les infections surviennent par contact avec ces fluides corporels ou avec des surfaces contaminées. Hors de l'hôte, les virus de la grippe peuvent rester infectieux pendant environ une semaine à la température corporelle humaine (37 °C), pendant plus de 30 jours à 0 °C et indéfiniment à très basses températures — comme les lacs du nord-est de la Sibérie. Ils peuvent être facilement inactivés par les désinfectants et les détergents[35].

Les virus se lient à une cellule par le biais d'interactions entre leur glycoprotéine d'hémagglutinine et les oses des acides sialiques situés à la surface des cellules épithéliales des poumons et de la gorge[36] (étape 1 sur le schéma du cycle de réplication). La cellule absorbe le virus par endocytose. Dans l'endosome acide, une partie de l'hémagglutinine fusionne l'enveloppe virale avec la membrane de la vacuole, libérant dans le cytoplasme les molécules d'ARN viral (ARNv), les protéines accessoires et l'ARN polymérase ARN-dépendante[37] (étape 2). Ces protéines et l'ARN viral forment un complexe transporté dans le noyau de la cellule, où l'ARN polymérase ARN-dépendante peut transcrire l'ARN complémentaire (ARNc), de sens positif[38] (étapes 3a et 3b). L'ARNc peut être exporté dans le cytoplasme pour y être traduit (étape 4), mais peut également demeurer dans le noyau. Les protéines virales nouvellement synthétisées peuvent être sécrétées par l'appareil de Golgi vers la surface de la cellule (dans le cas de la neuraminidase et de l'hémagglutinine, correspondant à l'étape 5b du schéma), soit être transportées vers le noyau pour s'y lier à l'ARNv et former de nouvelles particules de génome viral (étape 5a). Les autres protéines virales ont plusieurs actions dans la cellule hôte, notamment la dégradation de l'ARN messager cellulaire et l'utilisation des nucléotides libérés pour la synthèse de l'ARN viral, ce qui a également pour effet d'inhiber la traduction des ARN messagers de la cellule hôte[39].
Les ARNv de polarité négative qui forment le génome de futurs virus, l'ARN polymérase ARN-dépendante et les autres protéines virales sont assemblés en un virion. Les molécules d'hémagglutinine et de neuraminidase se regroupent en un renflement dans la membrane cellulaire. L'ARNv et les protéines structurelles virales quittent le noyau et s'insèrent dans le renflement membranaire. Le virus mature bourgeonne hors la cellule à l'intérieur d'une sphère de membrane phospholipidique de la cellule hôte, emportant l'hémagglutinine et la neuraminidase avec cette enveloppe membranaire[40]. Les virus adhèrent à la cellule par l'intermédiaire de l'hémagglutinine, de sorte que les virus matures se détachent de la cellule hôte où ils se sont formés une fois que leur neuraminidase a clivé les résidus d'acide sialique de la cellule hôte[36]. Cette dernière meurt après la libération des virions du virus grippal.
Les virus de la grippe, comme tous les Orthomyxoviridae, se répliquent donc dans le noyau cellulaire, caractéristique qu'ils partagent seulement avec les rétrovirus. Cela vient du fait qu'ils ne disposent pas de la machinerie enzymatique nécessaire à la production de leurs propres ARN messagers. Ils utilisent des ARN cellulaires comme amorce pour commencer la synthèse de l'ARN messager viral par un mécanisme appelé capture de coiffe (en)[41]. Parvenue dans le noyau, la protéine PB2 de l'ARN polymérase se lie à la coiffe de l'extrémité 5' d'un ARN pré-messager cellulaire. La protéine PA clive cet ARN près de son extrémité 5' et utilise ce fragment coiffé comme amorce pour transcrire le reste du génome de l'ARN viral en ARN messager viral[42]. Cette façon de procéder est nécessaire pour que l'ARN messager viral ait une coiffe afin de pouvoir être reconnu par un ribosome et être traduit en protéine.
L'ARN polymérase ARN-dépendante virale étant dépourvue de fonction de correction d'erreur, elle commet une erreur d'insertion de nucléotide toutes les 10 000 insertions environ, ce qui est à peu près la longueur du génome des virus de la grippe. Chaque nouveau virion contient ainsi statistiquement une mutation dans son génome[43]. La segmentation du génome en molécules d'ARN distinctes permet de mélanger (réassortir) les gènes lorsqu'au moins deux variétés de virus de la grippe infectent la même cellule. Il s'ensuit la formation d'un virus pourvu de propriétés différentes, pouvant par exemple permettre l'infection de nouveaux hôtes ou de se soustraire à l'immunité développée par l'hôte contre le précédent génome ; dans ce dernier cas, on parle de cassure antigénique[34].
Viabilité du virus et désinfection
Les virus de la grippe chez les mammifères ont tendance à être labiles, mais peuvent rester infectieux plusieurs heures dans le mucus[44]. Le virus de la grippe aviaire peut rester infectieux pendant 100 jours dans l'eau distillée à température ambiante, et pendant 200 jours à 17 °C. Il est inactivé plus rapidement dans le fumier mais peut demeurer infectieux pendant deux semaines dans la matière fécale. Les virus de la grippe aviaire peuvent rester infectieux indéfiniment lorsqu'ils sont congelés[44].
Les virus de la grippe sont sensibles à l'eau de javel, à l'éthanol à 70 %, aux aldéhydes, aux oxydants et aux ammoniums quaternaires. Ils sont inactivés par une température de 56 °C pendant au moins 60 minutes, ainsi que par un pH acide inférieur à 2[44].
Transmission des virus de la grippe
Tropisme et excrétion du virus de la grippe
Les virus de la grippe A possèdent un large spectre d’hôtes comprenant les oiseaux, les porcs, les chauves-souris, les vaches et les humains. Les virus de la grippe B constituent également un agent pathogène majeur chez l’homme, mais ne circulent pas chez d’autres espèces. Chez l’être humain, les virus A/H1N1, A/H3N2 et les virus de la grippe B provoquent des épidémies saisonnières chaque année.
Les principales cellules cibles des virus grippaux chez l’homme sont les cellules épithéliales tapissant le tractus respiratoire. Les voies respiratoires supérieures sont considérées comme le principal site d’infection, mais dans les cas sévères, lorsque des prélèvements des voies respiratoires inférieures sont réalisés, une réplication virale dans les poumons a été mise en évidence. Les virions nouvellement produits sont excrétés dans les fluides recouvrant la surface apicale de ces épithéliums et peuvent être dispersés vers de nouveaux sites du tractus respiratoire par l’action ciliaire, l’inhalation ou l’expiration.
Le virus est également libéré dans l’environnement par le nez ou la bouche, respectivement au sein des sécrétions respiratoires ou de la salive. Ces deux types de fluides constituent des matrices complexes contenant des sels, des enzymes, des glycoprotéines et d’autres microorganismes ; ces composants jouent un rôle important dans la détermination de la stabilité environnementale des virus de la grippe [45],[46].
Modes de transmission des virus respiratoires
Pas blanc ou noir Les définitions des aérosols et des gouttelettes ont évolué au cours des dernières décennies. Auparavant, on considérait que 5 µm constituait le seuil de taille séparant les aérosols des gouttelettes. Cependant, des gouttelettes expulsées d’environ 100 µm peuvent rester en suspension dans l’air et parcourir de longues distances, et ne correspondent donc pas à la définition classique des grosses gouttelettes[45].
De plus, la dynamique des aérosols expulsés est complexe : des études récentes ont montré que les aérosols expulsés peuvent changer de taille dans l’environnement, et que des aérosols plus petits peuvent se former à partir de gouttelettes plus grandes [47].La libération de sécrétions contenant des virus dans l’environnement crée un potentiel de transmission. Plusieurs modes de transmission sont possibles. Par exemple, les sécrétions contenant des virus peuvent être transférées directement à d’autres individus ou à des surfaces environnementales (fomites) par contact. La respiration, la parole, la toux et les éternuements libèrent tous des aérosols d’une large gamme de tailles, allant de quelques fractions de micromètre à 1 mm[47],[45]
Bien que l’importance relative de ces différents modes d’expulsion dans la transmission du virus de la grippe ne soit pas clairement établie, il est notable que, dans les modèles expérimentaux chez le furet, les virus de la grippe A transmis proviennent des voies respiratoires supérieures [48],[49],[50]. Les sécrétions peuvent être déposées dans les voies respiratoires supérieures d’un individu récepteur lorsqu’elles sont expulsées sous forme de projection composée de grosses gouttelettes (> 100 µm de diamètre) [45]. En raison de leur masse, ces gouttelettes se déposent rapidement hors de l’air et sont trop volumineuses pour être inhalées profondément dans les poumons.

La taille d’une particule expulsée, associée aux conditions de circulation de l’air, détermine la durée pendant laquelle elle reste en suspension dans l’air. Ainsi, les aérosols allant de moins d’un micromètre à environ 100 µm peuvent persister dans l’air après leur expulsion et parcourir de longues distances, créant une possibilité d’inhalation par un individu récepteur. Ces petites particules d’aérosol peuvent ensuite atteindre les voies respiratoires inférieures lorsqu’elles sont inhalées. Fait intéressant, la comparaison entre l’inoculation intranasale et l’inoculation par aérosol chez le furet suggère que le site anatomique de dépôt peut influencer à la fois la dose infectieuse et la probabilité de transmission ultérieure [51].
Les tentatives visant à différencier les modes de transmission se sont souvent appuyées sur la distance physique entre les individus infectés et les individus récepteurs comme indicateur indirect, en partant du principe que les aérosols peuvent parcourir de plus longues distances, tandis que la transmission par contact ou par projection de gouttelettes serait limitée à moins d’environ 2 mètres. Toutefois, ce raisonnement est problématique : tous les modes de transmission, y compris la transmission aérienne par des aérosols fins, peuvent se produire à courte distance. En outre, malgré leur capacité potentielle à se déplacer sur de longues distances, les aérosols peuvent ne pas être des vecteurs efficaces d’infection à longue portée. La dilution des aérosols lors de leur dispersion dans de grands volumes d’air est importante, et le nombre de virions qu’une seule particule d’aérosol peut contenir reste incertain. Les aérosols éloignés de leur source sont donc peu susceptibles d’apporter une dose de virus grippal suffisante pour provoquer une infection.
Outre la dilution, l’inactivation virale constitue probablement un facteur majeur. Les particules d’aérosol subissent des changements physiques au cours du temps, tels que la dessiccation et l’acidification [52] ; ces modifications peuvent endommager les virions contenus dans ces particules, de sorte que peu de virus restent viables dans un aérosol éloigné de sa source. L’absence de transmission à longue distance apporte donc peu d’éléments sur la capacité des aérosols à contribuer à la transmission à courte distance.
Bien qu’il soit difficile de démêler les modes de transmission responsables de la propagation dans chaque contexte humain, des données indiquent que tous les modes interviennent au cours d’une saison grippale. La transmission par aérosols des virus de la grippe a été documentée par des études observationnelles [53],[54], des essais contrôlés randomisés [55] et des expériences sur des modèles animaux [56],[57]. Par ailleurs, de grandes études épidémiologiques sur l’efficacité du lavage des mains suggèrent que la transmission par les fomites est responsable d’au moins une partie des infections grippales chaque année [58],[59]. Enfin, la transmission par contact direct au sein des foyers et des structures d’accueil de la petite enfance est considérée comme le principal moteur de la transmission des virus de la grippe B [59].
L'’efficacité relative de chaque mode de transmission varie probablement en fonction de conditions environnementales telles que l’humidité relative[60], la température[61] et la ventilation[62]. Ces facteurs modulent la dynamique des particules en suspension dans l’air, influençant la probabilité de transmission par aérosols, et peuvent également affecter la stabilité virale dans l’air et sur les surfaces [46],[63],[64]. Une complexité supplémentaire provient de la diversité des comportements humains susceptibles de favoriser la transmission et de la manière dont ces comportements varient selon le contexte social. Par exemple, les charges virales dans l’air peuvent varier en fonction de la fréquence et de l’intensité de la parole ; les surfaces des hôpitaux ou des structures de garde d’enfants peuvent être plus susceptibles d’héberger des virus infectieux que celles d’un supermarché ; et le contact direct est plus fréquent dans un cadre domestique que dans un environnement professionnel.
Prévention de la transmission de la grippe
Barrières naturelles
Conditions environnementales
La transmission en milieu intérieur se produit dans une plage de températures relativement étroite, mais l’humidité peut varier en fonction des températures extérieures, de la pression barométrique et des systèmes de régulation thermique intérieurs. Une faible humidité relative et/ou une basse température semble augmenter la transmission du virus de la grippe [60],[62],[65]. Des études épidémiologiques ont identifié une faible humidité absolue, plutôt que l’humidité relative ou la température, comme un facteur probable à l’origine des flambées saisonnières [66],[67],[68], mais ces résultats sont biaisés par le fait que l’humidité relative, la température et l’humidité absolue varient conjointement [46].
Composition des sécrétions respiratoires
Quel que soit le mode de transmission, les virus de la grippe sont transportés dans l’environnement au sein de matrices dérivées des sécrétions orales ou respiratoires. Les aérosols expulsés provenant de la cavité orale contiennent probablement une matrice de protéines et de sels très différente de celle des aérosols générés dans le tractus respiratoire [69],[70],[71] (Figure 2). De plus, les fluides issus de différentes régions des voies respiratoires supérieures et inférieures diffèrent entre eux, et l’état de maladie peut également modifier la composition des sécrétions [47],[71],[72]. Le liquide de surface des grandes voies aériennes humaines peut protéger les virus de la grippe de la dégradation induite par l’humidité [63]. La salive et plusieurs de ses composants ont également été rapportées comme inhibant l’infectiosité virale [73],[74].
Microbes commensaux et microenvironnement microbien
Un niveau supplémentaire de complexité provient de la présence d’une grande diversité d’espèces microbiennes au sein du tractus respiratoire et de la cavité orale. Les virus de la grippe transportés dans les sécrétions issues de ces sites sont susceptibles d’être accompagnés de bactéries et, potentiellement, d’autres virus (Figure 2). La modulation expérimentale du microbiome respiratoire de l’hôte a confirmé son potentiel à influencer la transmission des virus grippaux [75]. La co-infection par des bactéries pathogènes peut également modifier l’excrétion virale et l’efficacité de la transmission de la grippe [76]. La présence de certaines espèces bactériennes connues pour coloniser la cavité nasale peut altérer la persistance des virus dans l’environnement, offrant ainsi un mécanisme potentiel expliquant leur effet sur la transmission[75].
Contraintes physiques de l’environnement
Bien que les mécanismes conduisant à l’inactivation des virus dans les aérosols expulsés soient probablement multiples, l’une des hypothèses inclut les baisses de pH qui surviennent à mesure que les aérosols vieillissent [77]. En raison du mécanisme par lequel les virus de la grippe pénètrent dans les cellules — un processus régulé par le pH endosomal — les composants du virion grippal sont hautement sensibles à l’acidification et perdent leur infectiosité après exposition à des environnements acides [78]. Il est important de noter que le pH des aérosols peut varier en fonction de la composition de la matrice [77],[79], ce qui crée la possibilité que des aérosols générés à partir de sites anatomiques différents ou par des individus distincts soutiennent la persistance virale pendant des durées variables. La taille des gouttelettes constitue un second facteur clé : même lorsque la composition de la matrice est contrôlée expérimentalement, la décroissance du virus est plus rapide dans les aérosols que dans les gouttelettes de plus grande taille, et ce dans une large gamme de conditions environnementales [80]. Cette différence pourrait être liée aux propriétés physicochimiques spécifiques des aérosols expulsés [80].
Immunité préexistante contre la grippe
Un élément important à prendre en compte dans la circulation saisonnière des virus épidémiques, tels que la grippe, est la protection conférée par une immunité acquise antérieurement. Dès l’âge de cinq ans, la plupart des individus ont déjà été exposés à la grippe, et la mémoire immunitaire module la susceptibilité aux souches circulantes ou émergentes [81],[82]. Par exemple, lors de la pandémie de grippe H1N1 de 2009, les personnes nées avant 1950 étaient susceptibles de posséder une immunité préexistante les protégeant contre la souche pandémique [83],[84]. Cette protection est attribuée à des réponses immunitaires mémoires dirigées contre des souches ancestrales de H1N1 antigéniquement proches du virus pandémique H1N1 de 2009. En revanche, l’ampleur de la protection conférée par une infection antérieure par des souches antigéniquement éloignées, telles que celles possédant des sous-types distincts de HA et de NA, reste mal comprise.
L’impact de l’immunité préexistante sur la susceptibilité à l’infection à partir d’un individu donneur a été étudié à l’aide de modèles animaux [85]. L’impact de l’immunité préexistante dépend de l'intervalles de temps entre les expositions , même par un virus de la grippe A antigéniquement distinct, protégeait contre une réinfection par transmission [85]. Lorsqu’un modèle de transmission par aérosols a été utilisé, les furets récepteurs ayant une immunité contre le H1N1 n’ont pas été infectés par des animaux donneurs positifs pour le H3N2. Fait intéressant, l’effet inverse n’a pas été observé : l’immunité contre le H3N2 n’a pas réduit la susceptibilité à l’infection par le H1N1. Les effets protecteurs de l’immunité hétérosous-typique pourraient donc dépendre du sous-type ou de la souche virale. De manière surprenante, la barrière à l’infection par le H3N2 conférée par une immunité préalable au H1N1 pouvait être contournée dans un modèle de transmission par contact .
La barrière à l’infection conférée par une immunité préexistante pourrait modifier la dose virale nécessaire pour établir une infection réussie. Des travaux menés dans des modèles d’infection expérimentale chez l’homme dans les années 1960 ont suggéré que la dose infectieuse humaine 50 % (human infectious dose 50, HID₅₀) d’un virus de la grippe A de sous-type H2N2 était de 30 TCID₅₀ (dose infectieuse médiane en culture cellulaire) lorsque l’inoculum était administré par voie intranasale, ou de 3 TCID₅₀ lorsqu’il était administré sous forme d’aérosols [86].
Des études plus récentes utilisant des atomiseurs pour mimer une inoculation par aérosol ont toutefois montré qu’une dose infectieuse humaine beaucoup plus élevée, de l’ordre de 10⁶ TCID₅₀, est nécessaire pour les virus grippaux saisonniers humains H1N1 et H3N2 [87]. Cette différence marquée des valeurs de HID₅₀ pourrait être liée à des particularités des protocoles expérimentaux, mais pourrait également refléter des changements du niveau d’immunité dans la population, résultant de la vaccination ou d’infections naturelles antérieures.
Interventions non pharmaceutiques
Port de masque
Les humains expulsent des aérosols dont la taille varie largement (de moins de 1 µm à quelques millimètres de diamètre) et qui contiennent des génomes viraux. Par filtration selon la taille, les masques peuvent éliminer de l’air les aérosols que les individus inhaleraient normalement et empêcher le dépôt, dans le nez, de particules aéroportées de plus grande taille et de projections de gouttelettes. Ainsi, la filtration des aérosols peut réduire à la fois l’acquisition de l’infection par voie aérienne et la transmission directe à partir d’individus infectés. Avant 2019, des données indiquaient que le port du masque pouvait réduire la transmission « vers l’avant » des virus de la grippe, en particulier au sein des foyers [88], mais sa capacité à réduire le risque d’infection restait incertaine.
Les masques varient quant à leur efficacité relative pour filtrer des aérosols de différentes fractions de taille, ce qui a alimenté le débat sur la supériorité éventuelle de certains types de masques pour bloquer la transmission des virus respiratoires. Les masques N95 sont certifiés pour filtrer 95 % des aérosols de plus de 300 nm, tandis que les masques chirurgicaux filtrent environ 75 % des aérosols. Les masques chirurgicaux comme les masques N95 se sont révélés efficaces pour réduire l’émission d’aérosols contenant de l’ARN du virus de la grippe par des individus infectés [89], ce qui suggère soit que les aérosols de moins de 3 µm ne transportaient pas les génomes viraux dans la même mesure que les aérosols plus grands, soit que la collecte des aérosols n’était pas complète dans ces études. Les masques en tissu sont efficaces pour réduire les fractions d’aérosols de grande taille ; toutefois, les fibres libérées lors de leur utilisation rendent difficile une évaluation précise de leur efficacité de filtration [90]. Il est certain que les masques N95 sont supérieurs aux masques chirurgicaux, eux-mêmes supérieurs aux masques en tissu pour la filtration des petits aérosols. Néanmoins, tous semblent capables de réduire la quantité de virus libérée dans l’air et constituent probablement des moyens efficaces pour diminuer la transmission des virus de la grippe.
Limitation de l’exposition
Les risques spécifiques de transmission associés aux activités courantes ne sont généralement pas bien connus, ce qui constitue une lacune majeure dans notre compréhension de la transmission en milieu communautaire. Néanmoins, il est logique de penser que la réduction de la durée ou de la proximité de l’exposition à un individu infecté diminue les possibilités de transmission. La durée d’exposition généralement nécessaire pour qu’une transmission secondaire se produise n’est pas bien définie mais des études récentes menées sur des modèles animaux ont montré que des fenêtres d’exposition de 3 à 48 heures peuvent conduire à une transmission aérienne efficace des virus grippaux saisonniers [91],[92]. L’exposition au début de l’infection s'accompagne d'une augmentation des animaux récepteurs avec des animaux donneurs dans 3 à 5 premier jours [92],[93].
Ventilation
Le taux de renouvellement de l’air dans un espace intérieur peut influencer la concentration d’aérosols contenant des virus. La ventilation est depuis longtemps associée à une réduction de la transmission à longue distance des agents pathogènes aéroportés [94]. Des taux de ventilation élevés réduisent la persistance des aérosols contenant des virus dans un environnement donné, limitant ainsi les possibilités de propagation à longue distance, et diminuent la transmission dans les hôpitaux et les lieux de travail [95].
Lumière ultraviolette
De même, la lumière ultraviolette , en particulier les systèmes UV-C en partie supérieure de l’air ambiant, a été proposée comme méthode d’inactivation des virus présents dans les petits aérosols se déplaçant sur de longues distances [96],[97]. Des études animales récentes ont montré que l’inactivation par UV-C était efficace pour empêcher la transmission du SARS-CoV-2 entre des animaux séparés de plus d’un mètre [98]. Toutefois, la transmission des virus grippaux peut se produire à la fois à courte et à longue distance, et l’impact de la ventilation ou des systèmes UV-C en hauteur sur la transmission de la grippe chez l’homme demeure largement inconnu.
Interventions pharmaceutiques
Vaccins
L’efficacité vaccinale varie selon les saisons et se situe généralement entre 30 % et 50 % lorsque les souches vaccinales correspondent bien aux virus circulants. Comme l’indique cette efficacité relativement modeste, des infections de percée chez des individus vaccinés sont fréquemment observées. Néanmoins, les circonstances d’exposition qui permettent de surmonter la protection immunitaire restent mal comprises. Par exemple, il est possible que les individus immunisés présentent une susceptibilité réduite et nécessitent une dose virale plus élevée ou une voie d’exposition particulière pour être infectés. Par ailleurs, la transmission secondaire pourrait être moins probable à la suite d’infections de percée ; toutefois, le degré selon lequel la vaccination empêche la transmission ultérieure demeure incertain, en raison de niveaux faibles de traçage des contacts et de dépistage systématique des virus grippaux.
Malgré ces incertitudes, des taux élevés de vaccination sont considérés comme capables de réduire la charge communautaire de la grippe. Une étude rétrospective examinant l’impact de la vaccination antigrippale obligatoire chez les enfants d’âge scolaire au Japon a conclu que l’immunité collective dans ce groupe d’âge permettait d’éviter environ 40 000 décès par an [99]. Dans une autre étude centrée sur la transmission intrafamiliale, la vaccination d’un seul enfant par foyer a été estimée entraîner une réduction de 10 à 20 % des cas [100].
Dans les modèles animaux, les données sur la capacité de la vaccination à limiter la transmission du virus de la grippe sont rares et contradictoires [57],[101].
Thérapeutiques antivirales
Plusieurs composés de petite taille moléculaire ont été développés pour le traitement des virus grippaux. Ces composés ciblent différentes étapes du cycle de vie viral, telles que l’adsorption du virus, la fusion, la libération de l’acide nucléique, la réplication du génome viral et le bourgeonnement des virions. Cependant, l’accumulation de mutations chez les virus de la grippe aviaire augmente toujours la probabilité d’échappement immunitaire [102],[103]. Par conséquent, la mise à jour des médicaments antiviraux existants ne peut pas suivre le rythme de la variation continue des virus de la grippe.
Inhibiteurs de la protéine HA
Les inhibiteurs de la protéine HA bloquent l’adsorption ou la fusion virale et peuvent être divisés en inhibiteurs de HA1 et de HA2.
Les inhibiteurs de HA1, tels que le sulfate de dextrane et le DSA181 [104], empêchent la liaison de HA1 aux récepteurs de la surface cellulaire. Parallèlement, les inhibiteurs de HA2, comme l’arbidol[105] et le BMY-27709 [106], bloquent l’entrée du virus en empêchant la fusion membranaire médiée par HA2. Deux composés de petite taille moléculaire, MBX2329 et MBX2546, capables de se lier à la région de la tige du trimère HA et d’inhiber la fusion médiée par HA[107] ont été identifiés.
Protéases
Le processus de fusion du virus grippal dépend également de l’acidification endosomale et d’une série d’enzymes de l’hôte, telles que les protéases. Par conséquent, des inhibiteurs de ces enzymes de l’hôte ont également été développés comme médicaments anti-grippaux, notamment la bafilomycine A1[108] et l’aprotinine [109].
Inhibiteurs ciblant le canal protonique M2
Après la fusion membranaire, l’ARN viral pénètre dans la cellule hôte par le canal ionique M2. Des inhibiteurs de M2, tels que l’amantadine et la rimantadine, qui bloquent l’activité du canal ionique, ont été développés pour empêcher la libération du génome viral dans le cytoplasme.
Les inhibiteurs de M2 sont efficaces contre le virus de la grippe A, mais pas contre le virus de la grippe B, en raison de l’absence de protéine M2 chez ce dernier. Des doses prophylactiques d’adamantanes sont efficaces pour prévenir l’infection par le virus de la grippe chez l’homme [110]; toutefois, la résistance virale largement répandue à cette classe de médicaments en limite l’utilité [111]. La mutation S31N est la principale cause de la résistance aux inhibiteurs de M2, représentant ainsi 92 % des souches résistantes aux États-Unis [112]. En conséquence, les inhibiteurs de M2 ne sont actuellement plus recommandés pour le traitement.
Inhibiteur de la neuraminidase
La protéine neuraminidase est impliquée dans la maturation et la libération des virions et joue un rôle important dans la régulation de la liaison aux récepteurs et du bourgeonnement viral.
Les inhibiteurs de la neuraminidase, en particulier l’oseltamivir (également connu sous le nom de Tamiflu), sont largement prescrits pour le traitement de la grippe saisonnière et pandémique et sont autorisés pour un usage prophylactique aux États-Unis et ailleurs [113]. Les variants viraux résistants aux inhibiteurs de la NA ne sont pas actuellement très répandus chez l’homme, bien que des variants résistants soient fréquemment détectés chez les individus traités et aient été prédominants dans la lignée saisonnière H1N1 avant son extinction lors de la pandémie de 2009. Il est important de souligner que la prophylaxie post-exposition par oseltamivir chez les contacts étroits est efficace [114],[115], constituant un outil précieux pour prévenir la transmission. En revanche, le potentiel d’un traitement thérapeutique d’un patient index par oseltamivir pour empêcher la transmission est plus limité et dépend probablement fortement d’une initiation très précoce du traitement [116].
Toutefois, des mutations d’acides aminés dans la protéine NA, par exemple E119A, H274Y et N294S, entraînent généralement une résistance aux inhibiteurs de la NA [117],[118].
Inhibiteurs de la réplication de l’acide nucléique viral
Les inhibiteurs de la réplication de l’acide nucléique viral comprennent les inhibiteurs de PB2 [119](VX-787), les inhibiteurs de PA (flutamide[120] et baloxavir[121]), les inhibiteurs de l’ARN polymérase ARN-dépendante (favipiravir[122]) et les inhibiteurs de NP (nucléoline[123]).
Inhibiteur de l'endonucléase de la sous-unité acide de la polymérase virale
Le baloxavir n’a pas encore été largement évalué pour le blocage de la transmission, mais une étude suggère une efficacité comparable ou supérieure à celle des inhibiteurs de la neuraminidase [124]. Le traitement des animaux par baloxavir a supprimé l’excrétion virale chez les donneurs et réduit la transmission par contact des virus H1N1 et H3N2 après une exposition de 48 heures, comparativement à l’absence de traitement ou au traitement par oseltamivir [125].
Anticorps monoclonaux
Les anticorps monoclonaux ont été développés et suscitent de grands espoirs pour la prophylaxie post-exposition et le traitement clinique. Par exemple, un nouvel anticorps monoclonal humanisé, 8A5 [126], neutralise le H5N1 en se liant à deux types d’épitopes sur la protéine HA. Li et al. ont décrit un anticorps monoclonal chimérique, nommé C12H5, capable de neutraliser des souches représentatives de H1N1 circulant de 1991 à 2023, et même de neutraliser de manière croisée le H5N1 [127].