海馬 (脳)
From Wikipedia, the free encyclopedia
| 脳: 海馬 | |
|---|---|
 脳内での海馬の位置。赤で示した部分が海馬。 左は側面から見た図。右は正面から見た図。  | |
| 名称 | |
| 日本語 | 海馬 |
| 英語 | Hippocampus |
| 略号 | HIP |
| 関連構造 | |
| 上位構造 | 海馬体 |
| 画像 | |
| アナトモグラフィー | 三次元CG |
| 関連情報 | |
| IBVD | 体積(面積) |
| Brede Database | 階層関係、座標情報 |
| NeuroNames | 関連情報一覧 |
| NeuroLex | birnlex_721 |
| MeSH | Hippocampus |
海馬[1](かいば、英語: hippocampus)は、大脳辺縁系の一部である、海馬体の一部。特徴的な層構造を持ち、脳の記憶や空間学習能力に関わる脳の器官。
その他、虚血に対して非常に脆弱である[2]ことや、アルツハイマー病[3]における最初の病変部位としても知られており、最も研究の進んだ脳部位である。心理的ストレスを長期間受け続けるとコルチゾールの分泌により、海馬の神経細胞が破壊され、海馬が萎縮する。心的外傷後ストレス障害(PTSD)・うつ病の患者にはその萎縮が確認される[4]。βエンドルフィン(=脳内ホルモンの一つ)が分泌されたり、A10神経が活性化すると、海馬における長期記憶が増強する。
神経科学の分野では、海馬体の別の部位である歯状回と海馬をあわせて「海馬」と慣例的に呼ぶことが多い。本項では海馬のみならず、歯状回についてもあわせて言及する。
歯状回
海馬は細長い組織であるが、その長軸方向に沿って層構造が保存されているという特徴を持つ。そのため、海馬をどの位置で切断しても、その断面は後述の「ラット海馬の断面図」のようになる。前述のように、神経科学研究の世界ではこの断面図の状態を「海馬」と慣例的に呼ぶが、図中DGで示されている歯状回(dentate gyrus)と呼ばれる部位は海馬体の一部であって、正確には海馬の一部ではない。また、機能が関連している嗅内皮質についても多く言及しているが、嗅内皮質も海馬の一部ではない。
顆粒細胞(granule cell)と呼ばれる細胞の層を持ちその外側は分子層と呼ばれる。顆粒細胞は比較的小型の細胞であり、樹状突起を分子層側に、軸索を海馬のCA3領域の内側に向かって伸張させる。この軸索は苔状繊維とも呼ばれる。
海馬
主にCA1、CA2、CA3の各部位からなる。CAはアンモンの角を意味するフランス語、Cornet d'Ammonの略である。CA3野の錐体細胞は歯状回からのシナプス入力を受けている部位であり、CA2、CA1の錐体細胞層の内側に軸索を伸ばす。
※海馬の局所回路の詳細については、脳科学辞典の海馬を参照するとよい。
]
2
※遺伝子発現パターンに基づく海馬の解剖図については参考論文[5]を読むとよい。
嗅内皮質[6]から海馬へ神経投射(マウス)

図1の模式図の様に、嗅内皮質Ⅱ層の海細胞(Ocean Cell =neuron)は歯状回に投射し、島細胞(Island Cell = neuron;Wfs1を発現する)は海馬CA1領域のSL層に投射する。また、嗅内皮質Ⅲ層の神経細胞は海馬CA1領域のSM層に投射する。島細胞は、海馬CA1のSL層の抑制性神経細胞(GABA作動性)を介して、嗅内皮質Ⅲ層から海馬CA1への入力を抑制する。このように島細胞は嗅内皮質から海馬へのシグナルを抑制するので、学習中に島細胞を人工的に活性化させると条件付が成立しない。[7]
※ A→Bは、AからBへ投射するという意味とする。
海馬回路のアップデート

伝統的な海馬機能の見方は、長年合意されてきた情報の流れである嗅内皮質からの3シナプスループであり、それは歯状回やCA1,CA3を含みむもので、皮質へ戻っていく。この古典的回路は現在の我々が考える学習、記憶、空間ナビゲーションを着想させたが、海馬のその他の領域(CA2,fasciola cinereum,indusium griseum)や重要な視床下部からの入力を過小評価してきた。これらのまだよく研究されていない回路、伝統的でないネットワークが重要である。[8](2025)
CA3(海馬)への直接投射(マウス)

図3は、CA3領域に逆行性トレーサーを注入し、染色された神経細胞を3次元空間にプロットしたものである。この点1つ1つがCA3領域に直接投射するニューソンンの細胞体を示している。特に注目すべきなのが嗅内皮質の6b層(EC-6b)からCA3への投射が強いということである。EC-6bからCA3へは直接の興奮性投射がある。[9]
空間認識に関する海馬の機能
①場所細胞(Place Cell)について
海馬には、個体が特定の場所にいるときだけ特異的に活動する「場所細胞[10]」と呼ばれる神経細胞が存在する(基本事項については脳科学辞典参照)。また、海馬の場所細胞が経過時間を表現しており[11]、「時間細胞」と呼ばれることもある。
1)嗅内皮質からCA1への直接の入力を破壊すると、CA1の場所受容野の形成が障害される。[12]
2)CA1の浅層にある場所細胞は手掛かりが少ない環境下でより活動的で、海馬内の回路からの入力で発火頻度コード(位相コードよりも)をよりよく使うのに対し、CA1の深層の場所細胞は手掛かりが多い環境下でより活動的で、嗅内皮質からの入力で位相コードをよりよく使う。[13]
3)アルツハイマー病モデルマウスでは、場所細胞のリマッピングに障害が生じる。[14]
4)嗅内皮質Ⅲ層のCA1への入力を抑制すると、CA1の場所細胞の位相歳差(phase precession)が弱められ[15]、またCA1ニューロンの経験依存的な場所受容野形成を阻害する。[16]
5)歯状回顆粒細胞の入力や内側嗅内皮質の入力は、CA3における正常な位相歳差を起こすのに必要である。
6)歯状回ーCA3の回路について:ある部屋で場所細胞の場所受容野が形成されたときに、その発火パターンは、ネズミの、その部屋に入る入口によって変化する。この発火パターンの変化は場所細胞の発火頻度のリマッピングの一種で、CA1やCA3で認められる。この変化は、CA1において、歯状回ーCA3の回路を抑制することで起きなくなる。[17]
②速度細胞(Speed Cell)について
海馬には、個体の移動速度と発火頻度が相関する「速度細胞」が存在する。速度細胞は、介在ニューロンであることが多い。[18]
海馬と記憶の関係
※以下を読む前に、記憶についての基礎知識を脳科学辞典[19]で学んでください。
記憶痕跡(以下、エングラム[20])細胞をめぐる研究の展開
海馬と記憶が関わっているということは、様々な動物種における海馬の破壊実験によって明らかであった。20世紀初頭のカハール先生の研究以来、海馬については様々な形態的・機能的知識の蓄積が行われてきたが、2000年代に入り、急速なテクノロジーの進歩によって記憶研究のパラダイムシフトが起こった。どういうことかというと、神経回路機能と動物の行動(behavior)の関係を調べる研究が、相関関係を調べる科学から因果関係を調べる科学に変化したのである。この研究では基本的に「行動している最中の動物の脳」を調べる必要があるので、動物が死んだ後に切片を切って調べる方法では難しい。となると、生きたネズミに電極を刺して神経細胞の活動を調べることになる。しかし問題なのが、動物が行動しているときの神経細胞の活動は測定できるので行動と神経活動の「相関関係」は分かるが、その神経活動が実際にその行動の発生にどのくらい寄与しているかという「因果関係」は分からないのである。脳領域の細胞についての無差別な破壊や機能抑制をしたり、あるいは重要な遺伝子をノックアウトして動物の行動を調べる研究は行われていたが、それは脳の「領域レベル」や「遺伝子レベル」の因果的研究であって、「細胞レベル」つまり生命の基本単位としての因果的研究ではない。つまり、1つの神経細胞の発火が動物のbehaviorにもたらす役割が分からなかったのである。
そこで2000年代中ごろに登場したのが「光遺伝学(オプトジェネティクス)」[21][22]である。オプトジェネティクスは、行動している動物の脳の特定の神経細胞を発火させることができる。ある領野から領野に投射する神経細胞やある遺伝子を発現する神経細胞、また特定の学習記憶にかかわった細胞などを特異的に標識して発火させることができる。この技術によって、エングラム細胞を人為的に活動させることが可能となった。エングラム細胞とは、「記憶痕跡細胞」のことであり、「記憶の神経細胞単位の物理的実体」といってもよいだろう。2012年に、MITの利根川進博士は、マウスの歯状回にあるエングラム細胞を人工的に活動させ、記憶を想起させたという論文を発表した。
その論文[23]の概略を説明する。

部屋Bにいるマウスに電気ショックを与える。するとマウスは、部屋Bと電気ショックを関連付けてしまい、部屋Bを怖がるようになる。でも部屋Aについては関係ないので怖がらない。でも、もし部屋Bで電気ショックを受けたときに活動した歯状回の細胞(この学習におけるエングラム細胞)だけを人為的に活動させることができれば、部屋Aにおいても恐怖を想起させることができるかもしれない。そこで光遺伝学を使って、部屋Bで電気ショックを受けたときに活動した細胞(エングラム細胞)だけをいつでも操作できるようにしたのである。その仕掛けは後で説明する。実際にもともと恐怖と関係のない部屋Aにおいてこのエングラム細胞を人工的に活動させたところ、なんとマウスが恐怖反応を示したのである。
どのようにして、電気ショックのエングラム細胞だけを活動させられるようにするかを説明する(図5)。短く言うと、まずTet offシステム[24]を使用する(プロモータをc-fosにする)。c-fos(最初期遺伝子[25]の1つ:Immediate-early gene)という遺伝子は、神経細胞が活動すると発現するが、そのc-fosプロモータの下流にtTA(tetracycline transactivator)を付加したc-for-tTAマウスを用意する。すると神経細胞が活動するとtTAが発現する。さらにtTAと結合してプロモータの機能がonになるTRE(tetracycline-responsive element)プロモータとその下流に光遺伝学のチャネルロドプシン[26]をくっつけたベクターを導入する。すると、電気ショックで活動した細胞でc-fosプロモータが活性化し、tTAが発現する。(Doxがない状態では)tTAはTREに結合しチャネルロドプシンが発現する。チャネルロドプシンは細胞膜に到達し、光感受性の内向き陽イオンチャネルとして機能するようになる。その神経細胞に光をあてると、脱分極が生じ閾値を超えれば発火する。こうして、ある時間帯に活動した細胞だけを特異的に活動させる系を確立した。もしこの説明が分かりにくければ、原文(Susumu Tonegawa et al. Nature 2012)を和訳して読めばよい。
上記のようなセッティングをされたマウスを部屋Bにいれ、電気ショックを与えると、歯状回の、電気ショックに応答した細胞だけにチャネルロドプシンが発現する。つまり、歯状回のエングラム細胞がチャネルロドプシンで標識される。この状態で部屋Aにいるマウスの歯状回に光をあてると、恐怖と関係のない部屋Aで恐怖反応を起こすようになる。
リップル波(Sharp Wave Ripple)[27]
記憶の保持や将来の行動の設計には海馬のニューロンの同期した神経活動が重要である.このような同期した神経活動においては,海馬リップル波とよばれる脳波が発生する.[27]
海馬においてシナプスの多くは様々なタイプの神経活動に依存する可塑性を示す。もっとも典型的な可塑性は長期増強(long-term potentiation : LTP)[28]とよばれ、シナプスの結合強度が長期的に強まる現象である。長期増強は記憶が成立する上で不可欠である。しかし、人工的な刺激によりマウスの海馬に長期増強を引き起こし続けると記憶の機能が飽和することが報告されている。このことから、生じた長期増強を弱める機構が必要である。睡眠しているときには、海馬がや大脳皮質において自発的な長期抑圧(long-term depression : LTD)[29]が引き起こされる。これにより睡眠の経過とともにニューロン同士のつながりの強さが減弱し、記憶の機能の飽和を防ぐ。海馬においては、リップル波という脳波が長期抑圧を引き起こす。長期抑圧が引き起こされる結果、新奇の経験に関連しないニューロンの神経活動が選択的に抑制される。リップル波が不要なシナプスの結合強度を弱めることにより必要なシナプスのコントラストを強め、海馬から大脳皮質へと伝達される情報のシグナル/ノイズ比が上昇し、精度の高い情報処理が実現される。[30][31]
海馬のニューロンのごく一部は、脳波のシータ波と時には活動せず、リップル波のある時だけ活動するニューロン(Theta-OFF Ripple-ON:TOROニューロン)である。[32]
海馬と他領野の連携
※海馬の局所回路については脳科学辞典を参照

①海馬から前頭前野(PFC)への神経投射(マウス)
前頭前野(PFC)へ投射する腹側海馬(vHPC)の細胞は、浅層の細胞と深層の細胞に分けられる。浅層vHPC→PFC神経に比べて深層vHPC→PFC神経のほうが、樹状突起(最長の)の長さが長く、陽性電流を注入した後のバースト発火細胞の割合が多い。また光遺伝学的な実験が示すのは、浅層vHPC→PFC神経の発火がマウスの不安を低下させるのに対し、深層vHPC→PFC神経の発火はマウスの不安を強める。[33]
②学習における海馬と前部帯状回の連関(マウス)
CA1(海馬)と前部帯状皮質(AC)は機能的に関連している。
条件刺激と情動を伴う刺激を連合させる条件付学習を行ったとき、学習中と、想起中(条件刺激によって情動が励起される時)におけるCA1とACの神経活動を調べると、まずCA1において、情動をコードした神経細胞が出現する(情動を伴う条件刺激特異的に反応する細胞)。それに遅れてACで情動をコードした神経細胞が出現する。このことから作業仮説として(2022年現在)、外側嗅内皮質からCA1へのシグナル(単シナプスかどうかは不明)が重要な記憶保持回路で、ACからCA1へのシグナル(単シナプスかどうかは不明)が重要な記憶想起[34]回路であると考えられる。[35]

③前頭前野(PFC)⇔海馬の経路(直接的・間接的)
vHCPはPFC(眼窩前頭皮質:OFC,前辺縁皮質:PL、下辺縁皮質:ILを含む)の様々な領域に直接投射している。さらにPFCと海馬の間には、2つのPFC双方向の結合がある。1つはPLとILを起点に視床連結核(Re)を介してCA1に、もう1つはPLを起点にして嗅周野(perirhinal cortex:PRC)を介してCA1、外側嗅内野(LEC)を介してCA1やCA3に投射する。[36]
④海馬(CA1)からRetrosplenial Cortex(RSC)への投射
CA1のGABA作動性ニューロンはgranular RSC(RSCg)の1層(L1)に投射する。GABA作動性CA1→RSCgニューロンの長い軸索はRSCgにおいて、L5の錐体ニューロンを抑制する(L1にある尖端タフト樹状突起へのシナプスを介して)。化学遺伝学(Chemogenetic)的にGABA作動性CA1→RSCgニューロンを抑制すると、恐怖条件付け学習において恐怖の想起の段階における恐怖反応が増加する。GABA作動性CA1→RSCgニューロンからの抑制性軸索がRSCgのL1に入力し、遠位もしくは尖端タフト樹状突起に入力してL5の錐体ニューロンを制御する。海馬台のニューロンからの興奮性軸索は、深層にある、より近位の樹状突起に入力する。このように、GABA作動性の投射ニューロン(長い軸索を持った抑制性ニューロン)は脳の多くの領域で重要な役割をはたしている。[37]
⑤視床連結核(Re)からCA1への投射
Re→CA1ニューロンを抑制すると、恐怖条件付け[38]後の恐怖コンテキストでの恐怖反応が増加する。[39]
⑥腹側CA1から基底偏桃体(BA)への投射
腹側CA1→BAニューロンには腹側CA1→外側視床下部(LHA)ニューロンよりも多くの恐怖反応ニューロンが含まれる。
恐怖条件付の最中の腹側CA1のニューロンの恐怖反応は、恐怖記憶の形成(記銘)に必要である。[40]
発生
画像
 海馬の位置を様々な角度から眺めた動画。赤色で示す領域が海馬。(画像出典:Anatomography)
海馬の位置を様々な角度から眺めた動画。赤色で示す領域が海馬。(画像出典:Anatomography)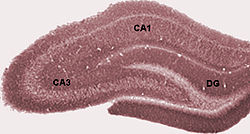 ラット海馬の断面図
ラット海馬の断面図 脳の海馬(左)とタツノオトシゴ、別名 海馬(右)。
脳の海馬(左)とタツノオトシゴ、別名 海馬(右)。


